
Écosystémique & coexistences créatrices
travail bibiographique en guise d'introduction à une écologie complexe & apports critiques issus de l'anthropologie
La présentation de certains des concepts suivants semble nécessaire afin de se départir de certaines approches encore trop réductionnistes au sein de l'écologie, comme au sein de la gestion des activités humaines en relation avec l'écosystème.
L'écologie et la biologie sont en effet à l'origine d'une rupture épistémologique importante qui n'est pas encore suffisamment prise en compte dans la réflexion à propos des relations au sein de l'éco-socio-système, et par conséquent souvent absente dans l'élaboration des actions entreprises afin de faire face aux problèmes auxquels nous sommes confrontés, comme celui de l'érosion de la biodiversité.
Cela a pour conséquence la pathologie du "contrôle et de la commande", ainsi nommée car elle est le symptôme d'une illusion quand à notre capacité de maîtrise des paramètres en jeu au sein des phénomènes naturels et sociaux, du fait de l'ignorance des phénomènes récursifs, des propriétés émergentes et des seuils propres aux systèmes auto-organisés.
Les concepts évoqués ici sont principalement issus du réseau de recherche www.resalliance.org, avec quelques compléments d'autres chercheurs dans le domaine de l'auto-organisation au sein des systèmes vivants. L'approche de ces socio-écosystèmes doit cependant être applatie et critiquée avec les apports de la sémiotique matérielle [https://gaiagraphie.hypotheses.org/1440].
Le dernière partie du document interroge la notion de co-gestion adaptative à partir des propositions d'Helen Verran de décoloniser les échanges de savoir.
Rond de fumée
"Un rond de fumée est littéralement et étymologiquement, introverti. Il tourne sans cesse sur lui-même comme un tore, un beignet, qui se meut autour de l’axe d’un cylindre imaginaire. C’est cette rotation qui permet à l’anneau de fumée d’exister en tant qu’entité distincte. Il ne s’agit, après tout que d’un peu d’air marqué d’un peu de fumée ; il est en fait de la même substance que son « environnement », mais c’est un phénomène durable, localisé, et qui, grâce à ce mouvement sur lui-même, forme une entité séparée. En un certain sens, l’anneau de fumée constitue un paradigme – très primitif et simplifié à l’extrême – de tous les systèmes récursifs qui contiennent les prémices de l’autoréférence ou encore si j’ose dire, de l’"égoité"."
(Bateson, 1986)
Systèmes complexes adaptatifs
"La relation entre structure et fonctionnement est l'un des fondamentaux de l'écologie. Les écosystèmes ainsi que la biosphère dans sa globalité, sont des exemples types de systèmes complexes adaptatifs, au sein desquels les propriétés macroscopiques telles que la structure trophique, les relations diversité-productivité, et les modalités de circulation des nutriments émergent des interactions entre les éléments, et peuvent en retour influencer le développement ultérieur de ces interactions."
(Levin S.A. 1998)
Résilience
La Resilience Alliance (www.resalliance.org) définit la résilience appliquée aux systèmes intégrant la nature et les êtres humains comme :
1. la quantité de perturbation qu’un système peut absorber et cependant se maintenir au sein du même état ou domaine d’attraction
2. le degré auquel le système est capable d’auto-organisation (versus manque d’organisation, ou organisation contrôlées par des facteurs extérieurs)
3. le degré auquel le système peut construire et développer sa capacité d’apprentissage et d’adaptation
(Carpenter et al. 2001a)
Le concept de résilience fait glisser les politiques de gestion depuis celles qui souhaitent contrôler le changement au sein des systèmes que l’on considère stables, vers la gestion de la capacité du socio-écosystème d’endosser, de s’adapter et de modeler le changement.
La gestion de la résilience renforce les possibilités de développement soutenable au sein d’environnements changeants ou le futur est imprédictible et la surprise attendue.
(Levin et al.1998, Holling 2001)
Au sein d’un système résilient, le changement recèle le potentiel de créer l’opportunité pour le développement, la nouveauté et l’innovation.
Au sein d’un système vulnérable, même de petits changements peuvent être dévastateurs.
La résilience fournit la capacité d’absorber les chocs tout en maintenant les fonctionnalités.
Quand un changement se produit, la résilience produit les éléments pour le renouvellement et la réorganisation.
(Gunderson and Holling 2002, Berkes et al. 2002).
La vulnérabilité est le côté sombre de la résilience : quand un système social ou écologique perd sa résilience, il devient vulnérable aux changements qui auparavant pouvaient être absorbés.
(Kasperson and Kasperson 2001a)
Cycle adaptatif
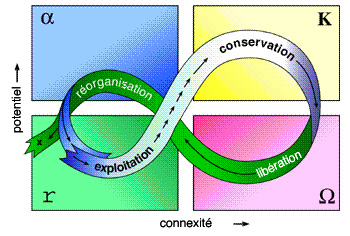
Les flèches montrent la vitesse de ce flux au sein du cycle, où les flèches courtes et proches indiquent une évolution lente, et une longue flèche un situation évoluant rapidement. Le cycle reflète le changement au sein de deux propriétés :
(1) axe Y - le potentiel inhérent à l'accumulation de ressources de biomasse et de nutriments ;
(2) axe X - le degré de connexité (connectance) parmi les variables de contrôle. Une faible connexité est associée à des éléments diffus faiblement connectés les uns aux autres, et dont le comportement est dominé par des relations externes, et affecté par la variabilité extérieure. Une forte connexité est issue d'éléments agrégés dont le comportement est dominé par les relations internes entre les agrégats, relations qui contrôlent ou atténuent l'influence de la variabilité exterieure.
La sortie du cycle à gauche de la figure suggère, l'étape où le potentiel peut s'échapper et où un saut vers un système moins productif et moins organisé est le plus probable.
(d'après Gunderson et Holling, 2002)
Au cours du temps, les structures et fonctions des systèmes changent du fait des dynamiques internes et des influences externes, résultant en quatre phases caractéristiques décrites par Holling pour la dynamique des systèmes écologiques :
•une phase de croissance (r) : accumulation lente de biomasse et de nutriments
•phase de conservation (K) : le système devient de plus en plus interconnecté, moins flexible, et plus vulnérable
•une période de libération de ressources limitées (Ω), après perturbation
•phase de réorganisation (α), menant ensuite à une autre phase de croissance au sein d’un cycle nouveau, avec une phase r similaire ou différente de la précédente
(Walker et al. 2006)
Stabilités alternatives
Tous les écosystèmes sont exposés à des changements graduels du climat, de la charge nutritive, de la fragmentation des habitats, ou de l’exploitation biologique.
La nature est habituellement supposée répondre aux changements graduels d’une manière régulière.
Cependant des bifurcations drastiques et soudaines vers un état contrasté peuvent interrompre ces changements réguliers, avec des conséquences sociales et économiques sérieuses. De même que des événements aléatoires tels que les tempêtes ou les vents peuvent induire ces bifurcations, de récentes études de prairies, de récifs coralliens, de forêts, de lacs et d’océans montrent que la perte de résilience prépare habituellement le chemin à la bifurcation vers un état alternatif (Scheffer et al. 2001). Les bifurcations entre états sont une des caractéristiques des systèmes complexes adaptatifs.
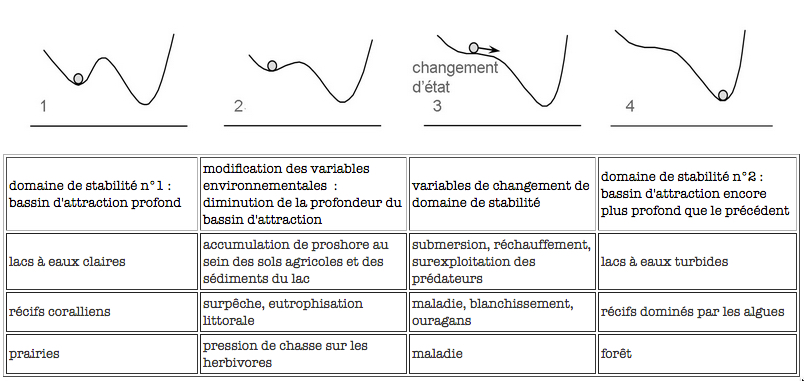
La diminution de profondeur du bassin d'attraction représente une perte de la résilience du système considéré, sous l’action de tel ou tel facteur environnemental, et se produit lentement. Le changement d'état du système (la bille qui glisse dans un nouveau bassin d'attraction), sera provoqué par un évènement soudain, et pourra être lui aussi très rapide.
Cycles emboités
Les modèles actuels concernant la relation entre la richesse spécifique et la stabilité modélisent implicitement les espèces et leurs fonctions écologiques à la même échelle ; cependant, les systèmes écologiques ne sont pas invariants d'échelles. Un corpus croissant de preuves empiriques, de théories et de modèles suggère que les structures et dynamiques écologiques sont principalement régulés par un petit ensemble de plantes, d'animaux et de processus abiotiques.
Ces processus opèrent à des échelles spatiales et des périodicités caractéristiques.
Les échelles petites et rapides sont dominées par des processus biophysiques qui contrôlent la physiologie et la morphologie végétales.
À l’échelle plus grande et plus lente de la dynamique des taches (patch dynamics), les compétitions interspécifiques des plantes pour les nutriments, la lumière et l’eau influencent les compositions et régénérations spécifiques locales.
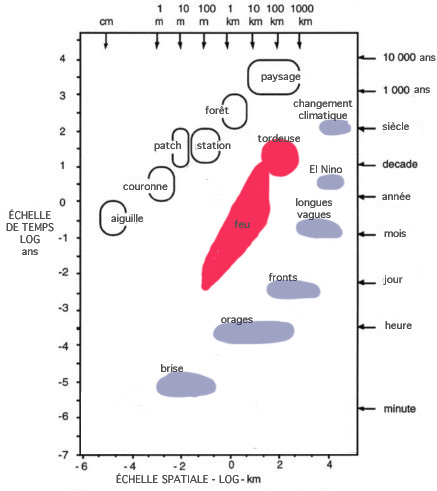
(d'après Peterson et al, 1998 x Holling et al. 2002)
À une encore plus grande échelle de la station forestière, les processus méso-échelle de feu, de tempêtes, d’attaque de phytophages et de consommation par les grands mammifères herbivores déterminent la structure et la dynamique successionelle depuis la dizaine de mètres au kilomètres, et des années aux décades.
À l’échelle du paysage la plus grande, le climat, la géomorphologie, et les processus biogéographiques modifient les structures et dynamiques écologiques depuis la centaine de kilomètres et au delà des millénaires.
Ces processus produisent de formes et sont à leur tour renforcés par ces formes ; cela signifie qu’ils sont auto-organisés.
(Peterson et al, 1998)
Panarchie
"C'est l'interaction entre les phénomènes à différentes échelles qui doit retenir notre attention. "
(Levin S.A. 1992)
Deux caractéristiques distinguent la représentation de la panarchie des représentations traditionnelles de hiérarchies emboitées.
La première considère l'importance du cycle adaptatif, et en particulier, la phase α comme moteur de la variété et le générateur de nouvelles expériences au sein de chaque niveau.
La seconde est la connexion entre les niveaux. Il y a potentiellement de multiles connections entre les phases d'un niveau et les phases d'un autre niveau. Mais deux d'entre elles sont déterminantes, et ont été intitulées “révolte“ et "mémoire“ dans la figure suivante où trois niveaux d'une hiérarchie sont représentés. Les connexions Révolte et Mémoire deviennent importantes aux périodes de changement du cycle adaptatif.
Quand un niveau de la panarchie entre dans sa phase Ω de destruction créatrice, cette destruction peut se propager au niveau supérieur plus lent en déclenchant une crise, particulièrement si ce niveau est dans une phase K, où la résilience est basse.
La flèche Révolte exprime la situation où des évènements rapides à une échelle inférieure bouleversent des processus lents à une échelle supérieure.
La flèche descendante appelée Mémoire indique le deuxième type d'interaction inter-échelles importante en période de changement et de renouvellement. Une fois qu'une catastrophe est déclenchée à un niveau, les opportunités et les contraintes pour le renouvellement du cycle sont fortement organisées par la phase K du prochain niveau, plus grand et plus lent. Après le feu au sein d'un écosystème, par exemple, les processus et les ressources accumulés à une échelle plus grande ralentissent la fuite des nutriments qui ont été mobilisés et libérés dans le sol. Et les options de renouvellement s'appuient sur la banque de graines, les structures physiques, et les espèces survivantes qui constituent l'héritage biologique accumulé pendant la croissance de la forêt. C'est comme si cette connexion s'appuyait sur la sagesse et l'expérience accumulée à la maturité, d'où le choix du mot Mémoire.
(Gunderson et Holling, 2002)

(modifié par C. Folke 2006 d'après Gunderson & Holling 2002)
Exemple d’une lande :
∞ mémoire : l’azote atmosphérique issu des nouveaux systèmes agricoles s’accumule dans les sols des landes et favorise les graminées dans leur compétition avec les éricacées
∞ révolte : le feu d’une prairie sèche en été se propage à l’échelle de la lande
∞ mémoire : le feu libère des nutriments stockés dans la matière organique et favorise les espèces capables de les mobiliser rapidement (fougères, graminées)
Une des caractéristiques essentielle de la panarchie, est qu’elle transforme les hiérarchies en structures dynamiques.
Les niveaux individuels présentent des propriétés non-linéaires à multiples états stables qui sont stabilisées ou déstabilisées par des connexions déterminantes entre les niveaux.
Il est certain que les niveaux lents et d’échelle plus vaste définissent les conditions au sein desquelles fonctionnent les niveaux plus rapides et d'échelle inférieure.
Ainsi une étendue forestière modère le climat et tamponne l’entendue des variations de tempétature que les espèces subissent.
Grumeaux
Sous l'action de forces centripètes et centriguges, les installations humaines de hameaux, villages, villes et agglomérations sont produites sous forme d'agrégats individualisés ainsi que de structures agglomérées de ces agrégats initiaux.
Par l'emploi du terme "grumeaux", il s'agit de qualifier non seulement de tels agrégats que l'on rencontre au sein des écosystèmes, comme les organismes qui en sont l'illustration évidente, mais aussi les associations végétales ou les écosystèmes eux-mêmes, cependant plus amorphes.
Mais en plus, il s'agit de qualifier aussi leurs attributs de taille, de vitesse et de fonctions qui sont distribués de manière grumeleuse. Ces attributs peuvent être des fluctuations périodiques, des tailles d'éléments à différentes échelles d'un paysage, des échelles de processus de décision d'humains et de non-humains, ou bien les attributs morphologiques et fonctionnels des animaux et des plantes.
Il existe deux raisons pour lesquelles une panarchie de paysage ou d'écosystème est susceptible de créer une forme grumeleuse.
La première est la nature discontinue des processus qui forment les éléments de la panarchie. Ce sont ceux qui créent une disjonction d'échelles parmi les variables structurantes clefs.
La seconde est la nature même du cycle adaptatif. Les phases du cycle sont distinctes et les changements entre ces phases sont abrupts, du fait des processus non-linéaires en jeu et d'un comportement multistable.
L'analyse des communautés animales au sein de paysages particuliers a révélé des structures multimodales, discontinues, parmi des attributs tels que la masse corporelle des animaux (Holling et al. 2002).
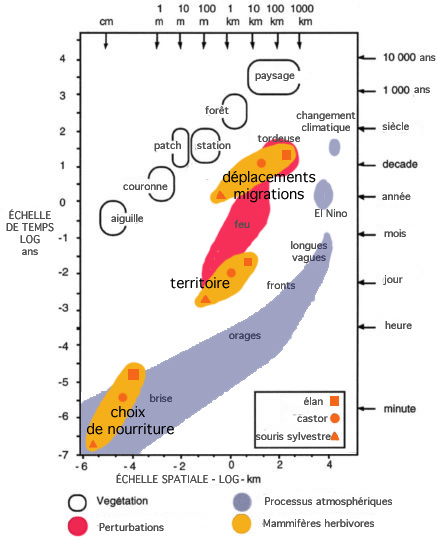
Les perturbations d'échelle médiane créent un lien intermédiaire entre les processus atmosphériques à l'échelle macro et les processus à l'échelle micro du paysage. Les échelles auxquelles la souris sylvestre, le castor et l'élan effectuent leur choix de nourriture, occupent un territoire et se dispersent afin de trouver ou rejoindre ceux-ci varient en fonction de leur taille corporelle (Peterson et al, 1998).
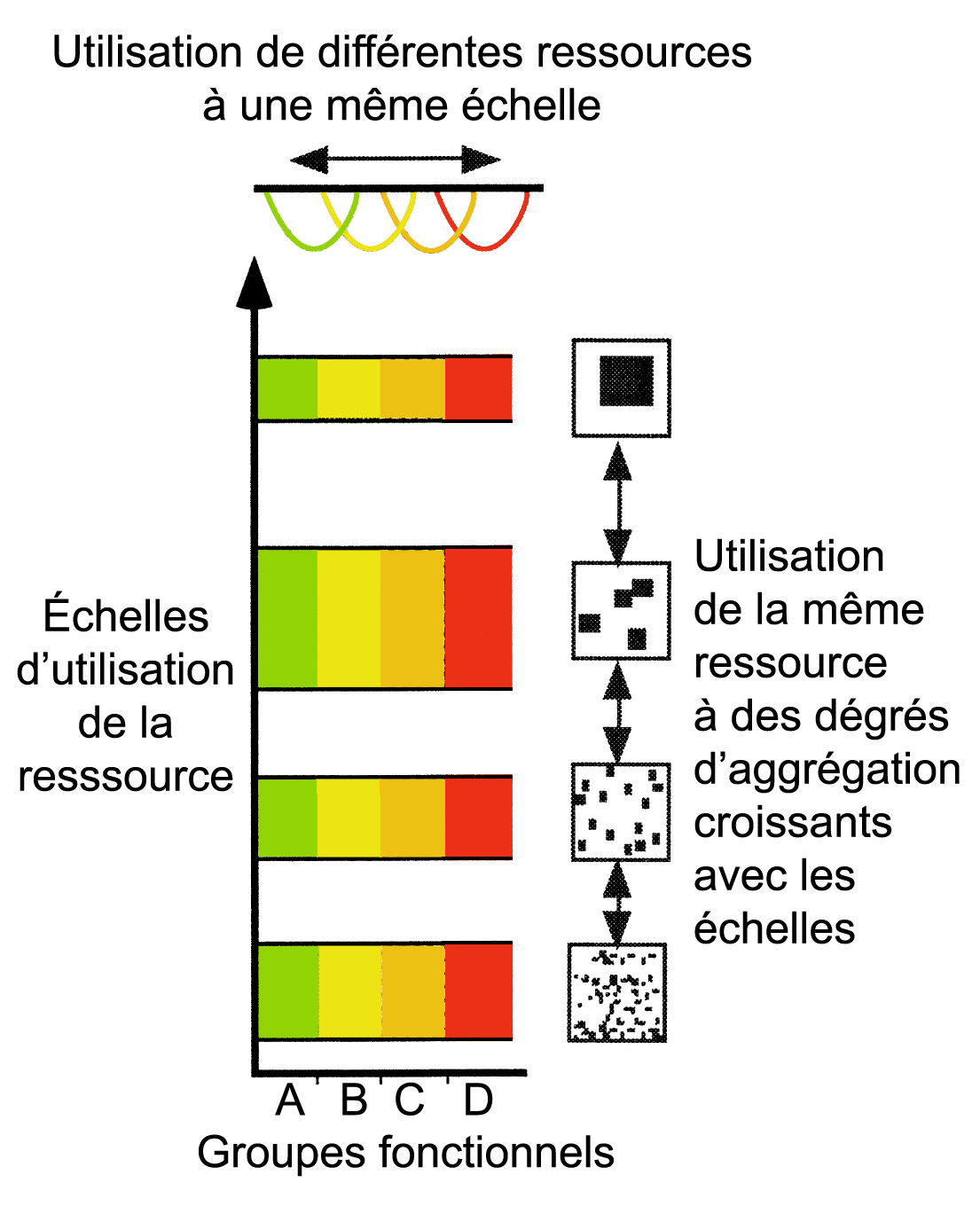
Des espèces différentes utilisent les ressources à différentes échelles spatiales et temporelles.
Les membres d'un groupe fonctionnel utilisent des resources similaires, mais les espèces qui opèrent à des échelles plus grandes ont besoin que ces ressources soient plus aggrégées qu e celles qui opèrent à des échelles plus petites.
Au sein d'une même échelle, la présence de différents groupes fonctionnels améliore le fonctionnement de l'écosystème, alors que la réplication d'une fonction à travers les niveaux d'échelle renforce le fonctionnement de celle-ci.
La combinaison de la diversité de fonctions écologiques à des échelles données, et la réplication d'une fonction à travers une diversité d'échelles produisent des fonctions écologiques résilientes (Peterson et al, 1998).
Les espèces peuvent être divisées en groupes fonctionnels déterminés par leur rôle écologique. Les espèces peuvent aussi être divisées en groupes selon les niveaux d'échelle particuliers de l'espace et du temps auxquelles elles exploitent une ressource. Les niveaux écologiques auxquelles les espèces opèrent correspondent souvent avec la masse corporelle moyenne, ce qui rend cette mesure utile afin de déterminer les échelles de perception et d'influence d'un animal (Peterson et al. 1998).
Un écosystème qui possède différentes niveaux d'échelle de structure écologique permet aux membres des guildes (ou des groupes fonctionnels) pluri-spécifiques de minimiser la compétition en utilisant les ressources disponibles à ces différents niveaux (Gunderson et al. 2002).
Cette réplication de la fonction à travers les échelles peut être observée à travers la dispersion des graines, depuis les fourmis, qui réalisent cette dispersion à faible distance au cours du trajet entre la ressource et la fourmilière, jusqu'à certains micromammifères ou oiseaux qui transportent ces graines une fois consommées, à travers leur tube digestif, à une distance plus grande. Cette dispersion à différentes échelles permet à la plante de persister malgré la variété des perturbations auxquelle elle est soumise (Gunderson et al. 2002).
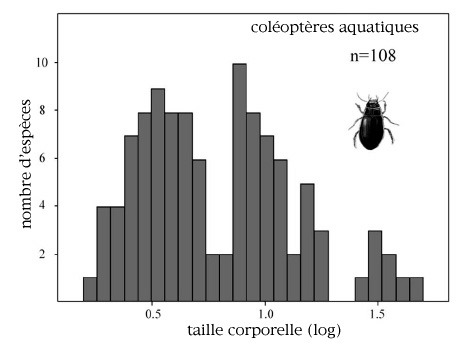
Bien que l'importance de la diversité fonctionnelle dans sa contribution aux services et bénéfices offerts par l'écosystème soit à souligner, la similarité fonctionnelle est un phénomène qui a lui aussi son importance, car il permet d'assurer la persistance des fonctionnalités écologiques en cas de changement des conditions environnementales. Cela signifie donc que la redondance générée du fait de cette similarité fonctionnelle est un facteur de résilience.
Ainsi, le déclin d'espèces dominantes est l'occasion pour des espèces mineures fonctionnellement équivalentes de se substituer à ces dernières.
Cette hypothèse a été testée à partir des fonctions écologiques liées au changement climatique global (Walker et al, 1999). À partir de l'identification de cinq attributs fonctionnels déterminants pour les échanges d'eau et de carbone (hauteur, biomasse, surface foliaire spécifique, longévité, qualité de la litière) de plantes herbacées de pâtures australiennes soumises à des pressions contrastées, l'hypothèse de résilience a été prouvée. Au sein de la communauté faiblement pâturée, les espèces dominantes étaient fonctionnellement plus dissemblables les unes des autres, et les espèces semblables d'un point de vue fonctionnel plus largement séparées dans les rangs d'abondance que ce qu'il est convenu d'attendre. Trois des quatre espèces mineures au sein des communautés faiblement pâturées, comme il était prédit par l'hypothèse, augmentèrent au sein de la communauté plus fortement pâturée.
La co-évolution au sein de communautés riches en espèces conduit à l'auto-organisation de structures grumeleuses d'espèces similaires sur le plan fonctionnel. Les effets structurants de la compétition sur les arrangements de la diversité spécifiques au sein de la nature doivent être regardés avec un regard nouveau. Il existe deux fenêtres d'opportunité permettant aux espèces de coexister : être suffisamment différent ou être suffisamment semblable. Le jeu de ces deux stratégies semble produire une structure auto-organisée de niches vers lesquelles les espèces convergent au cours de l'évolution (Scheffer & van Nes 2006).
Importance des grumeaux
Une fois que la structure de grumeaux et de creux est installée au sein d'une distribution, cela entraîne un ensemble complexe de variables associées entre elles. Cela détermine en partie le degré de résilience de la structure, et sa robustesse face aux modifications issues de sa gestion ou de de changements exogènes.
Comprendre ainsi la nature hiérarchisée des communautés animales et les ruptures d'échelle intrinsèques à ces communautés a permis une meilleure compréhension de la manière dont la résilience écologique et la durabilité sont issus de la diversité biologique.
Il existe deux grands types au sein de cette diversité biologique :
- celle qui affecte les fonctionnalité biologiques au sein d'un même niveau d'échelle (au sein d'un grumeau)
- celle qui affecte les fonctionnalités biologiques entre les niveaux d'échelle (entre grumeaux)
Ces deux types de diversité, que l'on peut nommer redondances imbriquées, produisent un renforcement des fonctionnalités qui est remarquablement robuste, et contribuent à la résilience et à la durabilité du système (Holling et al 2002).
Capacité adaptative
Selon Folke et al. (2003), la capacité adaptative est l’habileté avec laquelle un socio-écosystème se comporte avec de nouvelles situations sans perdre d’options pour le futur, et la résilience est la clef permettant cette capacité d’adaptation. Cependant cette notion de socio-écosystème doit être critiquée et applatie à l'aide d'une approche fine des modes d'assemblage et d'agentivité de ces collectifs hybrides [https://gaiagraphie.hypotheses.org/1440].
La capacité d’adaptation des systèmes écologiques est liée à la diversité génétique, la diversité biologique, et à l’hétérogénéité de la mosaïque paysagère.
Selon les chercheurs du réseau resaliance.org, l’existence d’institutions et de réseaux qui apprennent et conservent l’expérience ainsi que le savoir, apporte des solutions à la résolution des problèmes ainsi qu’un équilibre des pouvoirs parmi des groupes d’intérêts, et joue ainsi un rôle important dans la capacité adaptative (Scheffer et al. 2000, Berkes et al. 2002).
Folke et al. (2003) ont identifié et caractérisé quatre facteurs critiques qui interagissent à travers les échelles spatiales et temporelles et qui semblent être requises pour traiter avec la dynamique des ressources naturelles au cours des périodes de changement et de réorganisation :
• Apprendre à vivre avec le changement et l’incertitude ;
• Entretenir la diversité pour la réorganisation et le renouvellement ;
• Combiner plusieurs types de savoir pour l’apprentissage ;
• Créer des opportunités d’auto-organisation vers la soutenabilité socioécologique.
Apprendre avec le changement et l'incertitude
Provoquer la perturbation
De nombreuses communautés locales ont reconnu l’importance de la perturbation comme moyen de sécuriser les services de l’écosystème, et ont développé des pratiques de gestion qui miment les régimes de perturbation de la nature. Il existe des pratiques qui provoquent la destruction de l’écosystème en créant des impacts de perturbations à petite échelle.
En imitant ces perturbations naturelles à une échelle réduite, ces pratiques accélèrent le cycle de renouvellement local au sein de l’écosystème à plus large échelle, et permettent d’éviter l’accumulation de perturbation -la révolte - qui se propage au sein des niveaux d’échelle supérieurs et au delà au sein de la panarchie, l’ensemble hiérarchisé de cycles de renouvellement adaptatifs.
La gestion par le feu a été pratiquée largement par les sociétés traditionnelles en Australie et en Amérique du Nord afin d’ouvrir des clairières (prairies, marais), des corridors (chemins, affûts, franges herbeuses des cours d’eau et des lacs). Cela n’était pas utilisé à large échelle, mais plutôt avec une distribution en taches sur des habitats et des espèces ciblées. La gestion par le feu est aussi pratiquée dans la gestion contemporaine des forêts et des aires protégées (Folke et al. 2003).
Apprendre des crises
Une crise peut-être définie comme une large perturbation. Une surprise (désaccord qualitatif entre le comportement de l’écosystème et les prévisions) devient une crise quand il révèle un échec sans ambiguïté des actions et politiques de gestion.
Un exemple : la disparition de la ressources de morue à Terre-neuve avait été prévue par les pêcheurs de la zone littorale et quelques biologistes de terrain.
Le problème fut exacerbé par une trop grande confiance envers la science et la culture de l’évaluation quantitative des stocks du fait des modélisateurs des populations de l’Agence fédérale de gouvernement qui rétrospectivement admettent avoir mal évalué et mal interprété leurs données et précipitèrent une destruction du stock d’une magnitude sans précédent dans l’Atlantique Nord.
Cette destruction de la ressource aidera-elle l’agence de gestion à « répondre avec expérience » la prochaine fois qu’une crise analogue se produira ? Il n’y a pas de réponse claire à cette question car « répondre avec expérience » dépend d’une procédure d’apprentissage institutionnel basé sur des crises précédentes et une mémoire socio-écologique.
Les pratiques de provocation de la perturbation peuvent être le résultat d’un apprentissage institutionnel enregistré au sein de la mémoire des communautés locales, et peuvent aider à éviter des bifurcations qualitatives indésirables au sein des domaines de stabilité des systèmes naturels (Folke et al. 2003).
Prévoir l’imprévisible
Il existe de nombreuses pratiques locales de gestion et institutions associées qui évitent les crises à grande échelle. Au lieu de se débarrasser de la perturbation, l’existence d’incertitudes et de surprises et leur nature imprédictible est une part acceptée du développement, et les actions de gestion évoluent afin de faire face à leurs effets en répartissant les risques à travers la diversification à la fois des modes d’utilisation de la ressource et d’activités alternatives.
De nombreux groupes traditionnels d’agriculteurs conservent des variétés végétales à faible rendement comme assurance contre les risques climatiques et les maladies qui peuvent détruire de grandes quantités et variétés de végétaux. Ces stratégies sociales incluent l’investissement dans les cultures d’urgence, la polyculture comme stratégie contre les cyclones tropicaux, et la culture de plantes tolérante aux perturbations… Conserver des fragments de paysage permet aussi de servir de ressource supplétive face aux changements imprévus.
Ces pratiques sont le résultat de processus à long terme d’essais et d’erreurs permettant la réponse et l’adaptation socio-écologique à l’imprédictibilité environnementale (Folke et al. 2003).
Entretenir la diversité pour la réorganisation et le renouvellement
La diversité, en tant qu'élément de la résilience, offre aux systèmes socio-écologiques complexes la capacité de persister en cas de confrontation aux facteurs de changement. La gestion des systèmes complexes implique l'étalement des risques, la création d'espaces tampons et aussi de ne pas mettre tous ses oeufs dans le même panier. La diversité joue aussi un rôle au sein des processus de réorganisation et de renouvellement qui suivent une perturbation. C'est dans ce contexte que la mémoire sociale et écologique devient importante, car elle fournit un système d'expériences accumulées permettant de faire face au changement. C'est une ressource de créativité et de capacité adaptative.
(Folke et al. 2003)
Nourrir la mémoire écologique
Les espèces réalisent des fonctions clés au sein de l’écosystème.
Cependant, ce n’est pas le nombre d’espèces en tant que tel qui soutient un écosystème au sein d’un certain état stable ou domaine de stabilité, mais davantage l’existence de groupes d’espèces, ou groupes fonctionnels (prédateurs, herbivores, pollinisateurs, décomposeurs, modificateurs de flux hydriques, transporteurs de nutriments) avec des caractéristiques différentes, parfois partiellement redondantes.
Les espèces qui pourraient paraître redondantes et inutiles pour le fonctionnement de l’écosystème durant certaines étapes de son développement peuvent devenir d’une importance majeure pour régénérer et réorganiser le système après une perturbation et une destruction. D’autres ne le pourraient pas. De la même manière, les espèces redondantes connectent les habitats à travers leurs fonctions superposées au sein et entres les différentes échelles. La superposition (redondance) de diversité fonctionnelle augmente la variété des possibilités alternatives de modes et chemins de réorganisation qui suivent une perturbation et contribue à la résilience de l’écosystème.
La mémoire écologique est un élément important dans la superposition de la diversité fonctionnelle.
La mémoire écologique consiste en la composition et la distribution des organismes et leurs interactions dans l’espace et dans le temps, et inclu les expériences d’histoires de vies liées aux fluctuations de l’environnement.
La mémoire écologique consiste en au moins trois assemblages de base en interaction et leur diversité fonctionnelle superposée.
Le premier est l’héritage biologique, à travers les espèces et les formes qui persistent au sein d’une zone affectée après une perturbation, comme un arbre ayant survécu à un feu ou une graine qui requière le feu afin de stimuler sa germination.
Le second sont les liens mobiles, cad les espèces des groupes fonctionnels qui migrent entre les zones. Ces liens incluent les espèces qui se disséminent passivement d’une zone à l’autre, comme les larves à travers les courants ou les graines avec le vent, ou celles qui se déplacement activement entre les zones comme les poissons, les oiseaux, mammifères, et qui contribuent à la réorganisation de la zone perturbée.
Le troisième est constitué par les zones de soutien (zones sources) qui fournissent, au sein du paysage dont la zone perturbée fait partie, une diversité d’habitats pour les groupes fonctionnels à liens mobiles.
(Folke et al. 2003)
Combiner plusieurs types de savoir pour l’apprentissage
Le savoir et l’expérience des personnes au sujet de la gestion des écosystèmes contient des leçons sur la manière de répondre au changement et de nourrir la diversité. Ce troisième facteur souligne l’importance d’un tel savoir, son insertion au sein des institutions de gestion, et sa complémentarité avec les sciences et gestions conventionnelles des ressources. La compréhension scientifique des systèmes complexes adaptatifs peut être enrichie par des explorations des communautés locales et des sociétés traditionnelles possédant une expérience et une histoire continue de la gestion des écosystèmes. Il est aussi nécessaire, à partir du savoir relatif aux structures de la nature, de développer celui relatif au fonctionnement de la nature et à son rôle au sein de la résilience. Combiner différentes manière de connaître et d’apprendre permet à différents acteurs sociaux de travailler ensemble, même en contexte d’incertitude et d'information limitée.
Créer des opportunités d’auto-organisation
Selon Folke et ses collègues (2003), le quatrième facteur assemble les trois précédents dans le contexte d’auto-organisation. Soutenir la capacité d’interaction entre la diversité et la perturbation est un élément essentiel de l’auto-organisation. Le processus d’apprentissage est d’une importance primordiale pour permettre au socio-écosystème de construire sa résilience. Apprendre inclus l’utilisation du suivi permettant de générer et d’affiner le savoir et la compréhension écologique pour les institutions de gestion et les futures actions. De telles approches d’apprentissage sont présentes au sein de la co-gestion adaptative, un processus par lequel les arrangements institutionnels et les savoirs écologiques sont testés et révisés au sein d’un processus dynamique, permanent, et auto-organisé d’essais-erreur. Nous verrons cependant que cette ambition de construire un savoir partagé est largement critiquée par certaines anthropologues qui dénoncent le maintien d'un rapport colonial entre savoirs académiques et savoirs locaux et proposent comme remède de cultiver le dissensus.
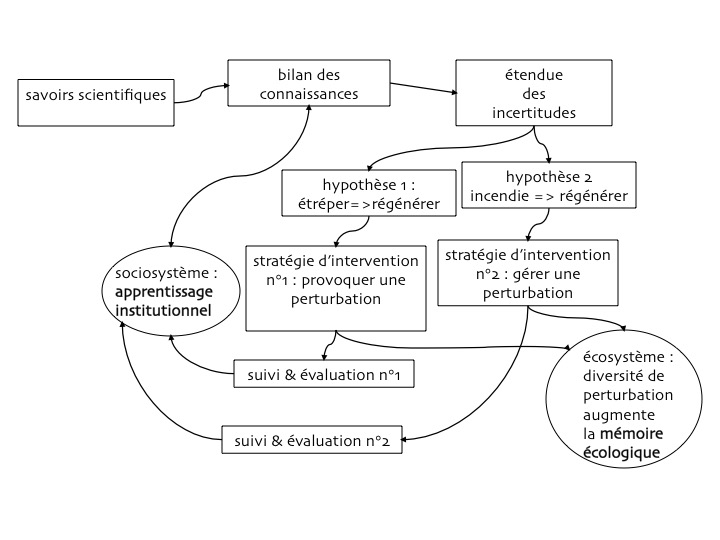
L’apprentissage institutionnel au sein du système social découle des stratégies de gestion mises en œuvre : en acceptant ou en provoquant des perturbations, cette gestion recherche la cohérence avec le cycle adaptatif, afin d’accroître la mémoire écologique
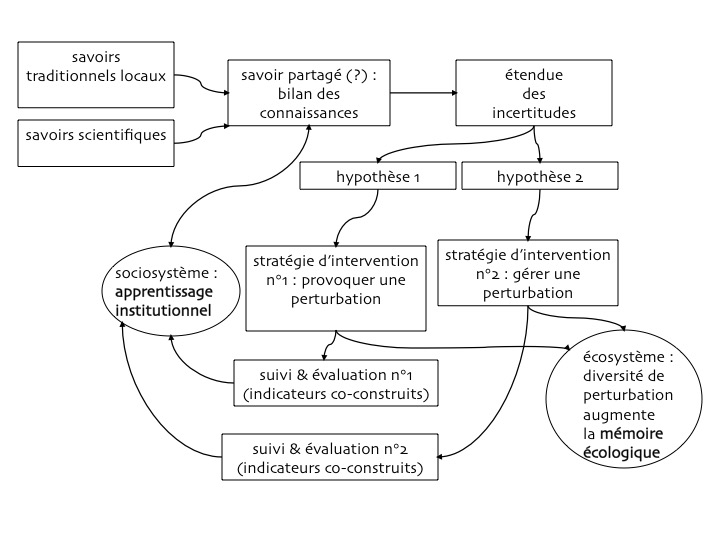
mêler les savoirs implique de co-construire les indicateurs, mais est-ce satisfaisant et suffisant ?
Décoloniser les échanges de savoirs

courtesy from Bonhams auction house
Chez les Yolngu de la Terre d’Arhnem, dans les Territoires du Nord de l’Australie, la logique n’est pas fondée sur l’emploi symbolique des nombres, selon notre base décimale liée à notre nombre de doigts, mais sur l’emploi d’un modèle relationnel fondé sur la parenté, nommé gurrutu, qui considère les relations entre trois générations au sein d’une famille : mari et femme, leurs 4 grands-parents et leurs 8 arrière-grands-parents, selon un ensemble de 16 paires de relations réciproques. Cet ensemble de relations forme un réseau de base, qui permet de positionner ensuite tous les êtres du cosmos Yolngu.
Dans la saison sèche, les Yolgnu mettent en oeuvre un schéma sophistiqué de brûlage de la végétation, selon un jeu de règles complexes et sacrées codées au sein des cycles de chants. Ces règles parlent de la direction des vents, la forme de la fumée, les conditions de sol, etc.
Les Yolgnu ne partagent pas notre conception d’un temps linéaire, et ne divisent pas le monde entre nature et culture. (Weirtheim, 1995)
Dans le cas de la plupart des aborigènes australiens, la relation d’appropriation mutuelle entre un territoire et sa population ne se réalise pas entre une surface délimitée et ceux qui y vivent, mais entre certains lieux et certaines personnes qui y sont reliés par les histoires de leurs clans et leur propres biographies (Ingold, 1987).
Ces lieux sacrés sont habités par les ancêtres totémiques, à qui des chants particuliers, des récits et des cérémonies sont associés. Ces lieux sont reliés par les sentiers des voyages des ancêtres. En d’autres termes, ces représentations spatiales sont à zéro dimension, (lieux, sites, emplacements), ou à une dimension (pistes, sentiers), plutôt qu’à deux dimension (surfaces délimitées), comme dans les sociétés agricoles. (Ingold, 1987)
Dans la carte suivante (Glowczewski, 2016), les pistes de Rêve Kangourou, Prune, ou Bâton à fouir peuvent se croiser, mais aussi se rencontrer sur des lieux sacrés qui sont alors associés à des obligations et à des droits rituels spécifiques, qui ne sont cependant pas figés mais évoluent avec les révélations oniriques. « Ces rêves réactualisaient dans les rituels les liens entre les Dreamings/ totems et les sites, créant ainsi une cartographie dynamique : les séquences de récits chantés qui relient en ligne des centaines de lieux totémiques nommés se sont ainsi renouvelées au cours du temps au rythme des morts, des naissances et de l’interprétation des rêves.” (Glowczewski, 2014)
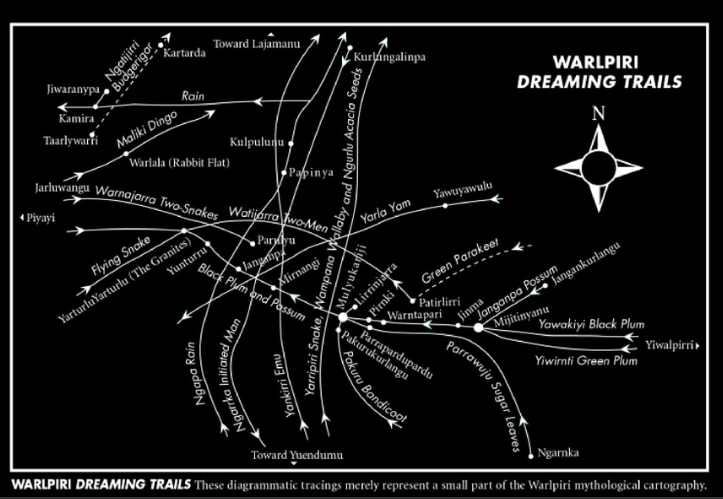
Sans reconnaissance de cette différence épistémologique et ontologique, aucune rencontre «post-coloniale» n’est possible. Lors d’un atelier avec des environnementalistes sur la résolution d’un problème commun à propos de savoir quand et comment faire les brûlages, Helen Verran (2002) insiste sur le fait qu’il est aussi important de participer à la manière dont le savoir est produit dans la pratique, à travers des rituels incorporés et hétérogènes. Les environnementalistes travaillent avec des quadrats, des mètres à ruban, des séries temporelles, et des relations inter-spécifiques.
Le scientifique est Linnéen, pour lui les deux plantes employées par les aborigènes pour allumer le feu sont des espèces différentes, c’est à dire des unités dans une taxonomie linéaire et hiérarchique, depuis l’espèce, au genre, à la famille, à l’ordre, etc., et elle ne se connectent dans une même unité qu’à la catégorie «ordre». Cette abstraction est familière de la méthode scientifique.
Les Yolngu travaillent avec des chants, des récits, des dances, et un système de relations de parenté récursif. L’ancien aborigène comprend les plantes comme un tout interrelié, ainsi que toute chose dans l’univers Yolngu. L’émergence des éléments de cet univers dépend du cycle nommé gurrutu. Dans ce cycle, les deux plantes qui génèrent le feu occupent la position originelle du cycle, leur unicité est une conséquence de la position singulière qu’elles occupent dans cet ensemble flou. Leurs savoirs sont différents parce que leurs rituels sont différents.
Nous devons prêter attention, de manière symétrique, à ces aspects pratiques de production du savoir, d’instant en instant. En agissant ainsi, il sera parfois possible de concevoir des rituels et formes de pratiques qui fonctionnent à travers les différences épistémologiques et ontologiques. (Law,2019)
Lors de l’atelier d’échange de savoirs sur la pratique des brûlages, une discussion s’engage entre un ancien aborigène et un scientifique sur les bois employés pour allumer le feu. Pour l’ancien, les arbres sont identiques, l’un est le petit-fils de l’autre.
Le scientifique dénie la similarité de ces bois, en employant les différences d’odeur pour les caractériser, ainsi que la position des feuilles, alternes chez l’un et opposées chez l’autre, mais ensemble ils finissent par reconnaître certaines ressemblances, comme un homme et une femme se ressemblent, mais sont différents.
Pour mener un engagement de bonne foi entre des traditions de savoir différentes, la tolérance est un bon début, mais insuffisant.
L’interrogation mutuelle, qui peut révéler à nous même nos propres traditions, autant qu’à l’autre, de manière symétrique, est ce qui est ici nécessaire (Verran, 2013:154) .
Une éthique du dissensus
Selon Helen Verran (2013:144), aborigènes et scientifiques ont besoin d’apprendre des manières de reconnaître les différences métaphysiques, de les valoriser, d’apprendre à refuser l’étape de réduction à des catégories partagées, et d’accepter que nous pouvons métaphysiquement ne pas être engagés dans un monde commun, et cultiver ainsi une posture décolonisée.
« [M]on travail comme intermédiaire culturel [...] aurait été plus fructueux si j’avais trouvé un moyen de maintenir et éventuellement d’exagérer cette irritation mutuelle en développant une technique qui fonctionne comme une forme de crispation épistémique.
En opposant des impulsions de bonne volonté bien intentionnées, organisées pour cultiver le consensus, de la part à la fois des scientifiques et des praticiens de la gestion de la terre Yolgnu (mais jamais de la part des autorités du savoir Yolgnu qui furent pratiquement sans compromis dans leurs présentations), quelques années plus tard j’ai vu que ce qui était nécessaire était une proposition de modalités acceptables pour les participants de manifester le dissensus publiquement et en tant que groupe, plutôt que séparément et presque secrètement, tel que cela s’est produit lors de la phase d’évaluation de l’atelier.
Je développe maintenant cet argument pour proposer que le dissensus explicite articulé comme élément de l’ici et maintenant de l’atelier, aurait révélé les feux comme menant à des objets singuliers de gouvernance, comme des évènements permettant de faire ensemble une différence.» (Verran, 2015)
Coexistences créatrices
«Les questions que se pose Verran sont «simples» : comment faire en sorte que des peuples inscrits dans des pratiques de savoir différentes «s’entendent » mutuellement, en particulier quand l’échappatoire simpliste du relativisme culturel n’est pas une option, ni politiquement, ni épistémologiquement, ni moralement ? Comment entretenir un savoir général dans des mondes post-coloniaux qui soient engagés à prendre les différences au sérieux ? Les réponses à ces questions ne peuvent être formulées qu’au travers de pratiques émergentes, c’est-à-dire au cours d’un travail précaire de terrain, rassemblant des capacités d’agir et des modes de vie discordants, qui puisse tenir compte à la fois de leur héritage historique disparate et de la possibilité, à peine possible mais absolument indispensable, de leur avenir commun. C’est dans ce sens que j’envisage les relations de partenaires.» (Haraway, 2018:30)
Poésie
Une raison pour laquelle la poésie est importante dans la découverte du monde est que, dans un poème, nous traçons un ensemble de relations depuis une diversité présente en nous-mêmes.
Ce niveau de diversité intérieur auquel nous n'avons pas accès normalement est exprimé à travers la poésie.
Nous pouvons ainsi nous offrir mutuellement, par la poésie, l'accès à un ensemble de relations avec l'autre et avec le monde dont nous ne sommes pas nous-mêmes conscients.
Grâce à cette cartographie de complexité à complexité, la poésie est un mode de connaissance du monde et de nous-mêmes.
(Bateson, 1972)

Bibliographie
Carpenter,S., B. Walker, J. M. Anderies, and N. Abel. 2001. From metaphor to measurement: Resilience of what to what? Ecosystems 4:765-781.
Collias, É. (2021, juin 19). Une recette de systémique plate [Billet]. coexister avec Gaïa, la terre vécue d’en bas. https://doi.org/10.58079/owvo
Bateson Gregory. In: Mary Catherine Bateson. Our Own Metaphor; a personal account of a conference on the effects of conscious purpose on human adaptaton. New York, Knopf, 1972.
Bateson Grégory, 1996. Cette histoire naturelle normative qu’on appelle l’épistémologie. In : Une unité sacrée : Quelques pas de plus vers une écologie de l’esprit. Seuil.
Berkes F and Folke, C. 2002. Back to the future: Ecosystem dynamics and local knowledge. In: Gunderson, LH and Holling CS (eds.) Panarchy: Understanding Transformations in Human and Natural Systems. Island Press, Washington DC.
Capra Fritjof , 2003. La Toile de la vie : Une nouvelle interprétation scientifique des systèmes vivants. Le Rocher.
Folke, C., J. Colding and F. Berkes. 2003. Synthesis : building resilience and adaptive capacity in social-ecological systems. Pages 352–387 in F. Berkes, J. Colding and C. Folke editors. Navigating social-ecological systems : building resilience for complexity and change. Cambridge University Press, Cambridge, UK. research interviewing. Sage Publications, Thousand Oaks, California, USA.
Folke, C., S. R. Carpenter, B. H. Walker, M. Scheffer, T. Elmqvist, L. H. Gunderson, and C. S. Holling. 2004. Regime shifts, resilience, and biodiversity in ecosystem management. Annual Review of Ecology, Evolution and Systematics 35:557-581.
Folke C. 2006. Resilience: The emergence of a perspective for social–ecological systems analyses. Global Environmental Change 16 (2006) 253–267.
Glowczewski, B., 2014. Rejouer les savoirs anthropologiques: de durkheim aux abo- rigènes. Horizontes Antropológicos, 20(41), 381-403.
Glowczewski, B., 2016. Desert Dreamers. U of Minnesota Press.
Gunderson, L.H., C.S. Holling, L. Pritchard Jr. and G.D. Peterson, 2002. "Resilience of large scale resource systems" Pages 3-20 in Resilience and behaviour of large-scale systems. L.H. Gunderson and L. Pritchard Jr., eds. Island Press, Washington, DC.
Gunderson LH and Holling CS. editors. 2002. Panarchy: Understanding Transformations in Human and Natural Systems. Island Press, Washington, DC.
Haraway, D., 2018. Manifeste des espèces compagnes. Climats.
Holling CS. 2001. Understanding the complexity of economic, ecological and social systems. Ecosystems 4: 390-405.
Holling, C.S. L.H. Gunderson and G.D. Peterson. 2002. "Sustainability and Panarchies" Pages 63-102 in Panarchy: understanding transformations in human and natural systems edited by L.H. Gunderson and C.S. Holling. Island Press, Washington, DC.
Ingold, T., 1987. The appropriation of nature - essays on human ecology and social relations. University of Iowa Press
Kasperson JX and Kasperson RE. editors. 2001a. Global Environmental Risk. United Nations University Press/Earthscan, London.
Law, J., 2019. Material semiotics. URL: www. heterogeneities. net/publications/ Law2019Material Semiotics. pdf.
Levin Simon A. 1992. The Problem of Pattern and Scale in Ecology: The Robert H. MacArthur Award Lecture. Ecology, Vol. 73, No. 6 (Dec., 1992), pp. 1943-1967.
Levin Simon A. 1998. Ecosystems and the Biosphere as Complex Adaptive Systems. Ecosystems 1: 431–436
Levin S, Barrett S, Aniyar S, Baumol W, Bliss C, Bolin B, Dasgupta P, Ehrlich P, Folke C, Gren I-M, Holling CS, Jansson AM, Jansson B-O, Martin D, Mäler K-G, Perrings C, Sheshinsky E. 1998. Resilience in natural and socioeconomic systems. Environment and Development Economics 3: 222-235.
Peterson, G., C. R. Allen, C. S. Holling. (1998). Ecosystem Resilience, Biodiversity, and Scale. Ecosystems 1: 6-18.
Robin A., 1958. Poésie non-traduite II. NRF Gallimard
Scheffer M, Brock W and Westley F. 2000. Mechanisms preventing optimum use of ecosystem services: An interdisciplinary theoretical analysis. Ecosystems 3:451-471.
Scheffer M, Carpenter SR, Foley J, Folke C and Walker B. 2001. Catastrophic shifts in ecosystems. Nature 413:591-696.
Scheffer and van Nes, 2006. Self-organized similarity, the evolutionary emergence of groups of similar species. PNAS April 18, 2006 vol. 103 no. 16 6203-6235.
Verran, H., 2002. A postcolonial moment in science studies: alternative ring regi- mes of environmental scientists and aboriginal landowners. Social Studies of Scien- ce, 32(5-6), 729-762.
Verran, H., 2013. Engagements between disparate knowledge traditions: Toward doing difference generatively and in good faith. Contested ecologies: Dialogues in the South on nature and knowledge, 141-161.
Wertheim Margaret, 1995. The Way of Logic. New Scientist, 2 December, pp. 38- 41.
Walker, B.H., Kinzig, A.P., Langridge, J., 1999. Plant attribute diversity, resilience, and ecosystem function: the nature and significance of dominant and minor species. Ecosystems 2, 95–113.